Материалы по биологии туркестанского змееяда (Circaetus gallicus heptneri; Falconiformes Accipitridae) на юго-западном Копетдаге
Букреев Сергей Анатольевич
Змееяд внесен в Красные Книги бывшего СССР и Туркменистана, поэтому любые современные сведения об этом виде представляют интерес. По материалам собственных исследований в 1984-1995 гг. и литературным данным характеризуется распространение и дается оценка численности Circaetus gallicus heptneri Dementiev в одном из слабо изученных участков его ареала – на Юго-Западном Копетдаге. Приводятся сведения по фенологии, гнездованию, успешности размножения, дается описание гнезд, яиц и постэмбрионального развития птенцов. Анализируются принятые и предлагаются дополнительные меры по охране вида в регионе.
Юго-Западный Копетдаг охватывает предгорные и горные области в пределах бассейнов рек Сумбар и Чандыр (кроме низкогорий на правобережье нижнего течения Сумбара) и характеризуется, по сравнению с другими участками Копетдага, относительно небольшими высотами (200-2000 м над ур. м.), более мягким, сухим субтропическим климатом и значительно лучшим развитием древесно-кустарниковой растительности (при общем сохранении доминирования открытых ландшафтов). Площадь туркменской части региона – около 4,1 тыс. км2.
С целью изучения биологии С. gallicus heptneri нами исследованы опубликованные в литературе сведения о змееяде на Юго-Западном Копетдаге; они очень незначительны и сводятся к указаниям на немногочисленные встречи одиночных птиц и пар этого вида (Зарудный, 1896; Ташлиев и др., 1965; Бельская и др., 1966; Л.С. Степанян, Е.Н. Степанян, 1972; Полозов, 1982, 1983; Мищенко, 1984), а также на находку единственного гнезда (Филатов, 1985). Кроме того, мы использовали неопубликованные материалы о встречах птиц, включая находки трех жилых гнезд, содержащиеся в отчетных книгах Летописи природы Сюнт-Хасардагского заповедника за 1979-1993 гг. (наблюдения С.И. Бурнашева, С.И. Забелина, В.Б. Зыкова, В.В. Кулибабы, В.В. Леоновича, B.C. Лукаревского, О.И. Роздиной, А.К. Сахарова, Н.В. Скалона, Т.А. Скалон, А.К. Филатова, Р.Ф. Штарева).
Собственные исследования проведены осенью 1984 г., летом и осенью 1986 г., во все сезоны 1987-1994 гг. и в марте-июне 1995 г. на территории всего Юго-Западного Копетдага. Пешими и автомобильными маршрутами обследовано около 2/3 площади региона, в том числе примерно 3/4 территорий, потенциально наиболее пригодных для обитания рассматриваемого вида. Нами найдено и описано 22 жилых и 16 старых гнезд, а также проведены многолетние (не менее 3 лет) наблюдения за пятью гнездящимися парами.
Распространение и численность
Змееяд на Юго-Западном Копетдаге – в целом довольно редкая гнездящаяся птица, однако в отдельных районах с особо благоприятными условиями его локальная численность может быть относительно высокой, и гнезда отдельных пар располагаются на расстоянии до 3 км друг от друга. На гнездовании вид связан с сухими, открытыми, изобилующими рептилиями угодьями. Именно наличие достаточного количества рептилий, а также их доступность (максимальная на открытых участках с низким, разреженным травянистым покровом) – основные экологические требования змееяда к своему местообитанию, определяющие в итоге пространственное распространение этого вида. Кроме того, для устройства гнезд он нуждается в каких-либо формах вертикального расчленения рельефа (крутые склоны, скалы, обрывы и т.п.) с разреженной древесно-кустарниковой растительностью, пусть даже самой низкорослой. Больших скал, сильно облесенных участков, узких и глубоких ущелий избегает, что опровергает утверждение Дементьева (1952) о том, что гнездовым биотопом змееяда должны являться “леса в Западном Копетдаге”, а также точку зрения Мищенко (1984а), причислившего змееяда в Копетдаге к обитателям зарослей лиственных деревьев нижнего пояса гор.
Основной район обитания в нашем регионе приурочен к опустыненным низкогорьям с абсолютными высотами до 500 м над ур.м., расположенным преимущественно в центральной и особенно в западной части Юго-Западного Копетдага и занимающим 1,65 тыс. км2, или около 40% его площади. Ландшафты среднегорий и нагорных плато имеют для вида второстепенное значение и его гнездование здесь – явление значительно более редкое. Самая высокая известная находка жилого гнезда – терраса правого берега р. Сумбар (около 800 м над ур.м.) между селами Тутлы-Кала и Коштемир (наблюдение B.C. Лукаревского).
Предпочитает селиться в укромных и редко посещаемых людьми местах, хотя некоторые пары, при отсутствии прямого беспокойства, быстро привыкают и могут спокойно относиться к регулярному выпасу и прогону вблизи гнезда домашнего скота.
Всего в настоящее время на территории Юго-Западного Копетдага известно 10 гнездовых участков змееяда с найденными жилыми гнездами. Еще в восьми точках мы встречали старые постройки, нередко здесь же в гнездовой сезон держались и сами птицы. Анализ всех известных встреч вида в репродуктивный период (рис. 1) позволяет очертить районы гнездования еще как минимум 21-22 пар. Таким образом, сейчас в рассматриваемом регионе выявлены места локализации примерно 40 гнездящихся пар змееяда, а с учетом необследованных и слабо обследованных территорий общая численность вида на Юго-Западном Копетдаге оценивается нами не менее, чем в 50-60 гнездящихся пар. Плотность населения в среднем по региону составляет 0,012-0,015 пар/км2, в том числе в оптимальных биотопах, расположенных на высоте ниже 500 м над ур.м., – 0,024-0,029, и в субоптимальных, расположенных выше 500 м над ур.м., – 0,004-0,005 пар/км2. Локальная плотность может быть несколько выше – например, 0,067 пар/км2 в предгорьях к северу и северо-западу от Кара-Калы, 0,05 пар/км2 в районе слияния Чандыра с долиной Варказ. Для сравнения укажем, что в Центральном Копетдаге (Ефименко, 1989, 1990, 1990а; Мищенко, 1986) и в Кугитанге (наши данные) змееяд является значительно более редким видом. Однако в более низкогорном Бадхызе его гнездовая плотность очень близка к плотности в нашем регионе и составляет 0,012-0,013 пар/км2 в среднем по всей территории Бадхызского заповедника и 0,03 пары/км2 в наиболее благоприятных местообитаниях (расчеты сделаны по данным Симакина, 1989).
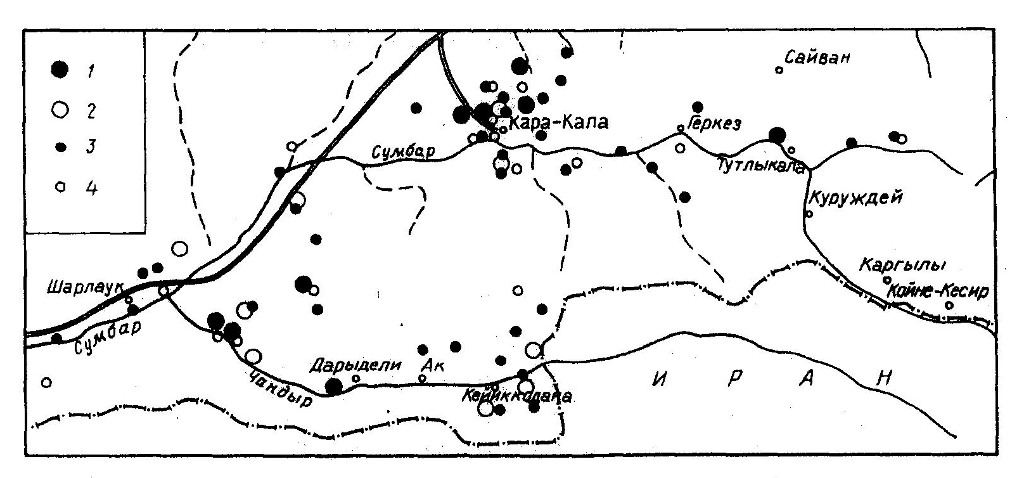{kind=link}
Межгодовые колебания численности гнездящихся птиц за весь период наблюдений нами не отмечены, что свидетельствует об относительной стабильности кормовой базы этого вида.
Характер и сроки пребывания
Перелетный вид. Весной прилетает в последних числах марта – начале апреля, т.е. когда уже выходит из зимовки основная часть рептилий (Скалон, 1982; Щербак и др., 1986). В качестве примера можно привести следующие даты первых встреч в различные годы: между 1 и 5.IV 1980, 8.IV 1981, 2.IV 1985, 9.IV 1986, 3.IV 1987, 23.III 1988, 18.III 1989, 22.III 1990, 25.III 1991, 3.IV 1992, 22.III 1995 гг., в среднем за эти 11 лет – 29 марта ± 2 дня. Осенью основная часть птиц отлетает уже в сентябре, но отдельные запоздалые особи могут наблюдаться и в первой половине октября: нам известно 11 таких октябрьских встреч, в том числе самые поздние даты в разные годы – 8.Х 1984, 6.Х 1986, 10.Х 1987, 12.Х 1988, 5.Х 1990, 5.Х 1991, 8.Х 1992, 1.Х 1993 гг. Таким образом, время пребывания вида в нашем регионе составляет 181-204 (193 ± 3; n = 6) дня.
Хорошо выраженного транзитного пролета змееядов через Юго-Западный Копетдаг мы не наблюдали, их общая численность в периоды пролета не отличается от летней. Скоплений, даже временных (например, в богатых кормом местах) этот вид никогда не образует. Чаще всего отмечаются одиночки или пары птиц. В период послегнездовья и осеннего пролета иногда попадаются тройки, в которых одна из птиц всегда молодая (чуть более темная), что согласуется с существующим мнением (Корелов, 1962) о возможности отлета на зимовку семейными группами.
Связь с территорией, гнездовой участок
Гнездовые участки отдельных пар очень постоянны. Нам известны случаи, когда птицы не покидали свой участок даже после гибели гнезда или одного из партнеров. Все пары, за которыми проводились многолетние наблюдения, ежегодно гнездились на одних и тех же участках. Привязанность к гнездам выражена значительно слабее. Иногда в одном и том же гнезде птицы поселяются 2 года подряд, но чаще они каждый год гнездятся в разных местах. Например, одна из наблюдавшихся нами пар (рис. 2) в течение 7 лет гнездования сменила семь гнездовых построек (как минимум три из которых существовали уже в первый сезон наблюдения за этой парой), другая пара за 6 лет гнездилась в пяти местах, третья – за 3 года сменила два гнезда. Расстояние, на которое новое гнездо удалено от прошлогоднего, составляет от 100 м до 1 км. Если под гнездовым участком понимать ту территорию, в пределах которой расположены все гнездовые постройки пары (и которая поэтому охраняется от других представителей своего вида), то его площадь колеблется от 0,2 до 4 км2. Охотничий участок всегда совмещен с гнездовым, но во много раз больше его по площади.
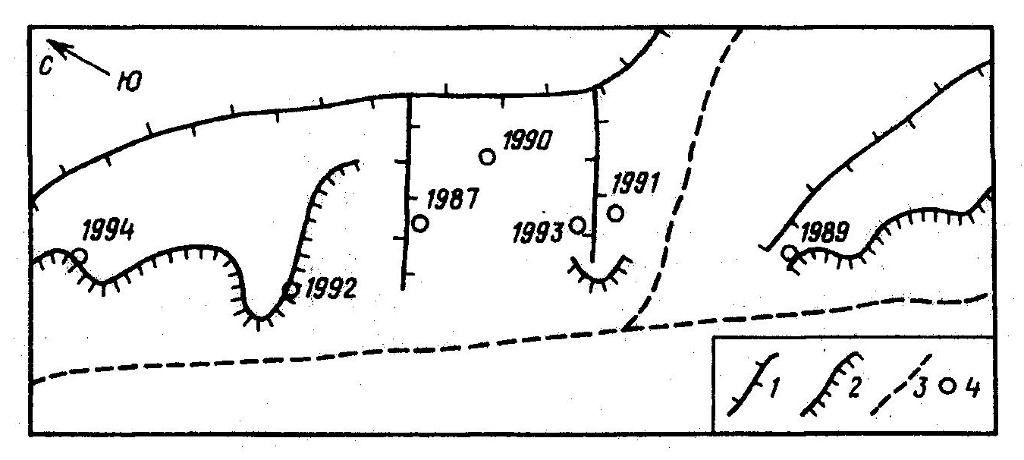{kind=link}
Считается (Корелов, 1962), что змееяды избегают соседства других хищных птиц. Наши наблюдения в целом подтверждают этот факт, но дело здесь, видимо, не в антагонизме, а в довольно специфическом выборе змееядом мест для устройства гнезда, которые далеко не всегда отвечают требованиям других хищных птиц. Скорее, к другим крупным птицам змееяд относится нейтрально и достаточно спокойно мирится с тем, что они поселяются в пределах его гнездового участка (такие случаи отмечены нами для балобана Falco cherrug, курганника Buteo rufinus, белоголового сипа Gyps fulvus, стервятника Neophron percnopterus, обыкновенной пустельги Falco tinnunculus, ворона Corvus corax). Один раз жилые гнезда змееяда и белоголового сипа располагались всего в 80 м друг от друга.
Расположение и устройство гнезда
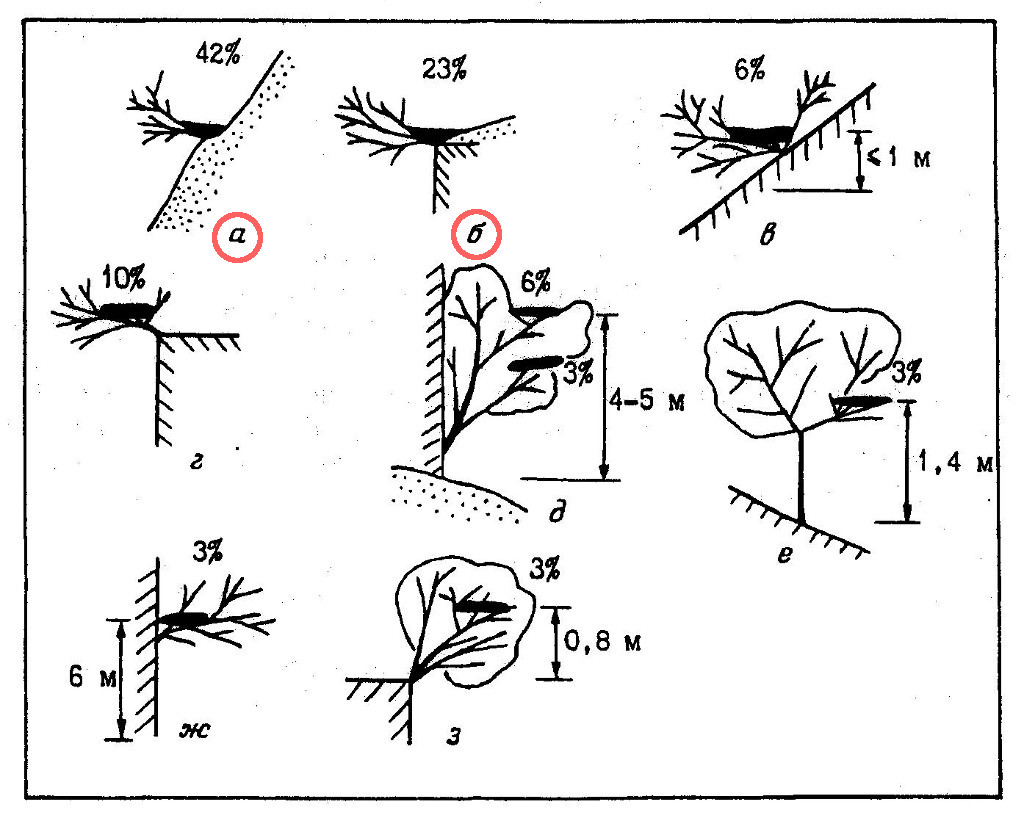
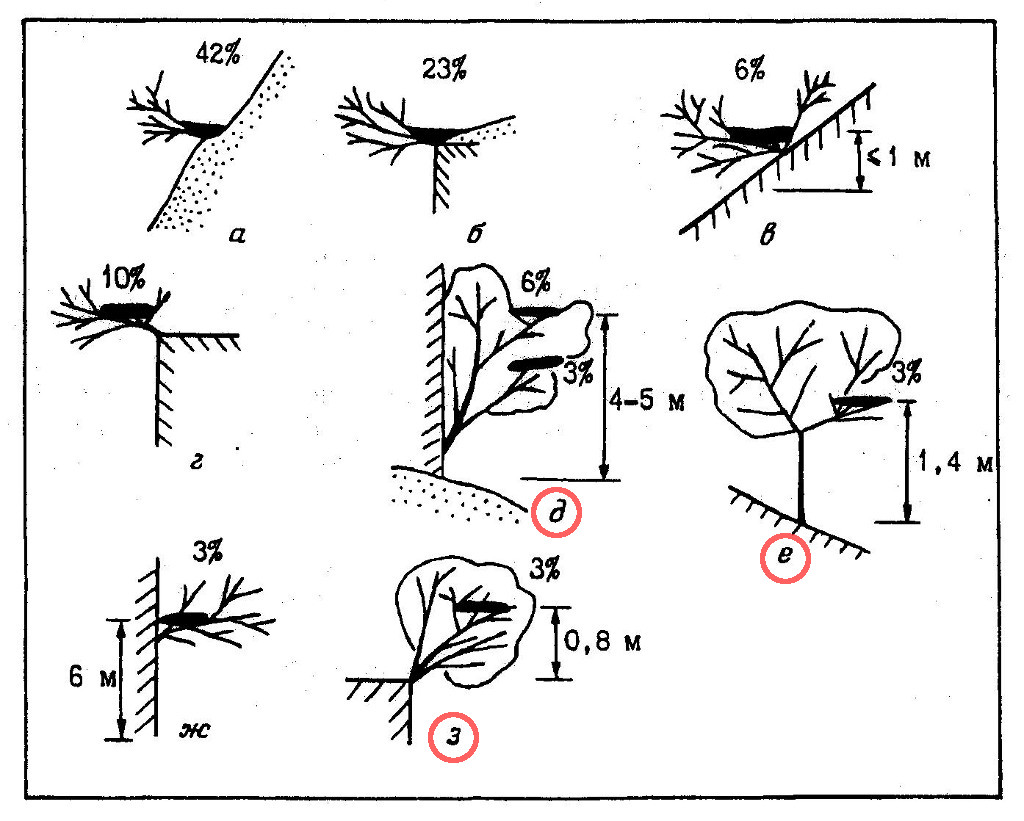
Несмотря на привязанность к сухим ландшафтам, змееяды при размещении гнезд сохраняют ярко выраженную дендрофильность. Все известные нам гнездовые постройки этого вида так или иначе были связаны с деревьями и кустарниками. Различные типы устройства гнезд и частота их встречаемости показаны на рис. 3. В подавляющем большинстве случаев (65%) дендрофильность носит номинальный и, так сказать, редуцированный характер (рис. 3, а, б). Гнезда при этом устраиваются на крутых (45-50° и более) голых склонах холмов, на такой же крутизны скалах и у верхних кромок обрывов, но всегда в основании небольших кустов. Нередко ствол и крона куста в качестве опоры для гнезда используются чисто символически, и основная часть постройки (до 2/3 общей площади), что включая и весь лоток, лежит прямо на поверхности земли. При гнездовании на склонах холмов птицы предпочитают устраивать гнезда в их средней части, на расстоянии 20-25 м от подножья, реже – в верхней части, в 7-10 м от вершины. Определенного предпочтения в выборе экспозиций склонов и обрывов не отмечено.
{kind=link}
{kind=link}
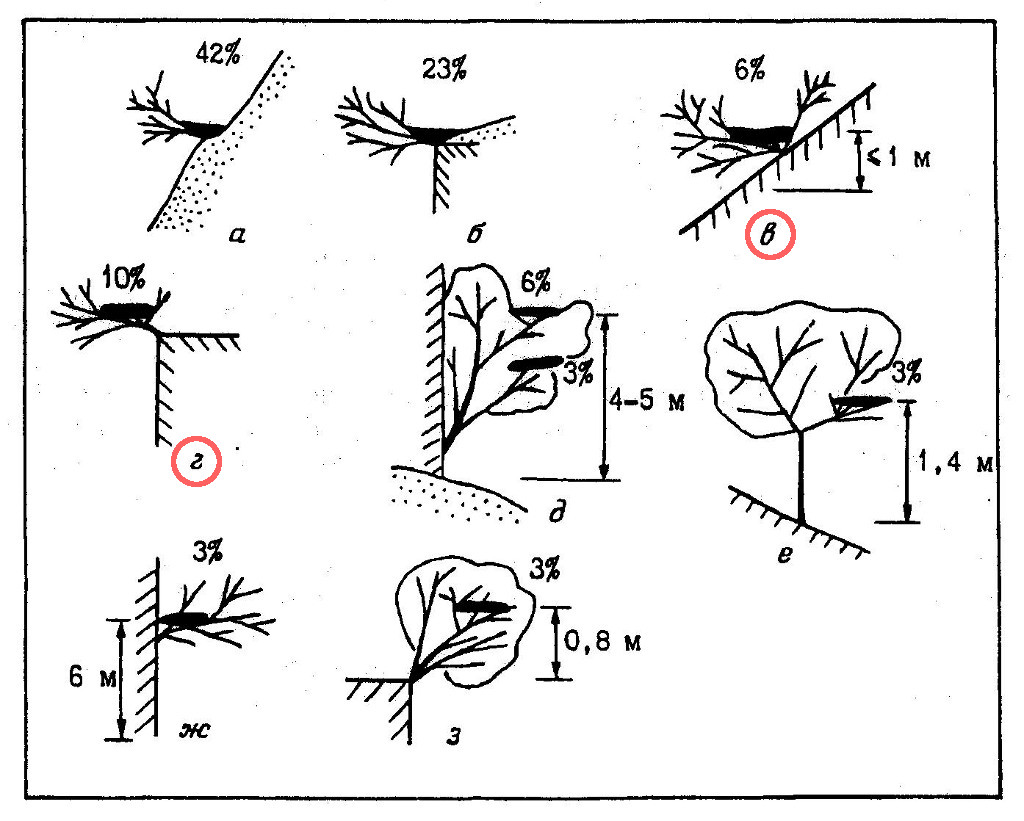
В 19% случаев гнезда змееядов целиком располагались на низкорослых кустарниках (на высоте всего 20-40 см от их основания), растущих в аналогичных описанным выше местах (рис. 3, в, г), а также непосредственно на обрывах (рис. 3, ж). Гнездование в кронах невысоких деревьев отмечено в 15% случаев (рис. 3, д, е, з). Как уже отмечалось выше, слишком больших скал и густых лесов змееяды избегают, гнездование их на высокоствольных деревьях нам также не известно.
{kind=link}
{kind=link}
{kind=link}
Из древесно-кустарниковых пород при устройстве гнезд предпочтение отдает парнолистнику Zygophyllum atriplicoides (46% из 26 гнездовых построек; типы устройства гнезд а, б), используются также жостер Rhamnus coriacea (23%; типы а-г, ж), инжир Ficus carica (15%; г, д, з), каркас Celtis caucasica (8%; д, е), арча Juniperus turcomanika (4%; в) и мелкоплодная вишня Cerasus microcarpa (4%; г).
Гнезда почти всегда располагаются очень доступно как для человека, так и для четвероногих хищников, что является самым уязвимым местом в охране вида и служит одной из основных причин гибели насиживающих птиц, кладок и птенцов. Всего лишь одна из всех известных нам гнездовых построек располагалась в средней части скалистого обрыва на относительно недоступной высоте (рис. 3, ж).
Чужих гнезд змееяды не занимают и всегда строят их сами. Рыхлые и нередко просвечивающие стенки гнезда (особенно используемого первый раз) сооружаются из тонких веток; лоток выстилается грубыми частями различных травянистых растений, а также мелкими веточками полукустарников и кустарников – почти всегда со свежими зелеными листьями, которые обновляются в течение всего периода насиживания; в гнезде и вокруг него всегда имеется небольшое количество собственных перьев и пушинок, по которым легко узнать жилые гнезда даже издали. Общий диаметр гнездовой постройки от 50 х 40 до 80 х 70 см, в среднем (n = 17) 59 ± 3 см; высота – 10-30 (22 ± 2; n = 15) см. Лоток плоский и неглубокий, его диаметр от 23 х 18 до 30 х 30 см, в среднем (n = 15) 26 ± 1 см и глубина 3,5-10,0 (6,6 ± 0,6; n = 9) см.
Репродуктивный цикл
Формирование брачных пар, занятие будущих гнездовых участков и даже попытки гнездостроения могут происходить у змееядов уже за 1-2 года до наступления полной половой зрелости. Так, одна из наблюдавшихся нами пар впервые появилась на новом участке, где змееяды в предыдущие годы никогда не отмечались, весной 1993 г. В этом, а также в следующем году эта пара демонстрировала брачное поведение и в одном и том же месте начинала строить гнездо, но затем без видимых причин прекращала это занятие, хотя и продолжала еще довольно длительное время держаться в пределах данного участка. И лишь в 1995 г. в том же самом гнезде у этой пары впервые появилась кладка.
К размножению приступают ежегодно. На гнездовых участках появляются сразу же после прилета, часто уже парой.
Ритуал совершаемых в воздухе брачных игр довольно разнообразный, в литературе описываются различные элементы репертуара и у туркестанского подвида (Абдусалямов, 1971; Корелов, 1962; Симакин, 1989). Мы несколько раз наблюдали парные полеты самца и самки, во время которых птицы с дистанцией всего в несколько метров друг от друга синхронно парят и планируют, как правило, на довольно большой высоте и совершают различные воздушные эволюции. В одном случае, например, пара полого планировала с сильно вытянутыми вперед шеями и согнутыми почти под прямым углом, и в то же время широко расправленными на концах крыльями и издавала при этом не очень громкие крики. Иногда самец совершает одиночный демонстрационный полет. Один раз мы наблюдали парящую птицу, издававшую протяжный, мелодичный свист, отдаленно напоминающий песню черного дрозда.
В целом же нужно отметить, что змееяды по природе довольно молчаливы и даже в брачный сезон характеризуются слабой вокализационной активностью. Будучи спугнутыми с гнезда, они также предпочитают молча парить на большой высоте или же молча в отдалении наблюдать за происходящим, сидя на удобной вершине. И только когда в гнезде маленький птенец, самка может довольно активно окрикивать агрессора с ближайших холмов или летать при этом низко у него над головой, имитируя пикирование. Тревожный крик самки змееяда у гнезда представляет собой серию несколько протяжных, односложных, чередующихся звуков, полный структурный рисунок которых можно передать как “кхе, кьйо, кьйо, кьйо, кьйо, кьйо – пауза около 4 с – кьйо, кьйо, … кьйо (до 15 раз, с нарастанием интенсивности позывок в середине фразы)” (рис. 4).
{kind=link}
Игривость у змееяда сохраняется в течение гнездового периода довольно долго, вплоть до вылупления птенцов, а по мнению некоторых (Корелов, 1962), – даже до вылета молодых. Воздушные игры над гнездом происходят в вечернее время. Вот один из таких случаев, наблюдавшийся нами 4.VI 1987. В 9 час. вечера обе птицы парили над гнездом, в котором находился маленький пуховик. Самец часто совершал следующие пируэты: плавно поднявшись на большую высоту, он затем круто пикировал вниз, тут же снова вздымал вверх, достигнув исходной высоты, на короткое время переходил в горизонтальный полет, затем пируэт повторялся и так 5-6 раз подряд. При выходе вверх из воздушной “ямы” птица сильно вытягивала вперед голову.
К ремонту и строительству гнезд змееяды приступают практически сразу же после появления на гнездовом участке. Гнезда в стадии строительства или ремонта мы находили 21.III, 4 и 6.IV 1989 г., 23.III и 1.IV 1995 г., 12.IV 1991, 16.IV 1992 и 19.IV 1990 гг. Гнездо строят обе птицы 7-10 дней (Симакин, 1989; наши данные). Самка начинает ночевать в гнезде еще до начала яйцекладки. С момента окончания строительства гнезда до откладки яйца проходит от двух до трех недель.
Гнезда с яйцами регистрировались нами со 2-й декады апреля по начало июня, самая ранняя дата – 11.IV 1989 (свежее яйцо), самая поздняя – 6.VI 1987 (насиженное яйцо). Судя по всему, у основной части птиц яйцекладка происходит во второй половине апреля, реже – в первой половине этого месяца и в начале мая. Единственная известная нам указанная выше июньская кладка была снесена новой самкой, которую привлек на свой гнездовой участок самец, потерявший в самом начале репродуктивного периода первую партнершу.
После гибели кладки к ее возобновлению в текущем сезоне змееяды, видимо, приступают очень редко. По крайней мере в двух случаях, когда птицы потеряли яйца слабой и средней насиженности, их гнезд с новыми повторными кладками, несмотря на специальные поиски, мы найти не смогли. Однако, Симакин (1989) по материалам из Бадхыза описывает один факт возобновления кладки после гибели слабо насиженного яйца, причем в том же самом гнезде. На возможность этого указывает и Дементьев (1951) для номинативного подвида C.g. gallicus.
Кладка во всех известных нам случаях (n = 19) состояла из одного яйца грязновато-белого цвета, форма яйца округлая, скорлупа толстая, Матовая и шероховатая. Длина яиц 72,1-78,5 (74,4 ± 0,4; п = 14) мм, поперечный диаметр 53,2-59,0 (57,2 ± 0,5; n = 14) мм; вес свежих и слабо насиженных яиц 110,3-138,5 (129,8 ± 5,2; n = 5) г, сильно насиженных – 101-127 (117,5 ± 8,2; n = 3) г.
В насиживании кладки принимают участие оба партнера, но самец проводит в гнезде значительно меньше времени, чем самка, сменяя последнюю лишь на короткий период кормежки. Сведения о длительности инкубационного периода в литературе очень противоречивы. Называются разные сроки от 30 (Корелов, 1962), 35-37 (Аверин, 1981; Воiнственський, 1984; Гавриленко, 1929; Никифоров и др., 1989; Штегман, 1937) до 42-48 дней (Атаев, 1974; Сухинин, 1957; Ивановский, 1992; Шерешевский, 1931; Makatsch, 1974). Собственных прямых наблюдений за продолжительностью инкубации мы не проводили, косвенная же оценка дает срок около 40 дней, так как птенцы в гнездах вылупляются в конце мая – начале июня (25.V-4.VI, по результатам прямых наблюдений и расчетным данным для девяти обнаруженных с пуховыми и только начавшими оперяться птенцами гнезд).
Насиживающая самка при появлении возле гнезда человека затаивается, плотно “вжимаясь” в лоток, и сохраняет полную неподвижность, лишь изредка слегка приподнимая для осмотра голову. В случае сильно насиженной кладки или недавно вылупившихся птенцов она может подпустить к себе на расстояние до 10 м. Слетев с гнезда, старается незаметно скрыться из вида низко над землей и в присутствии человека попыток вернуться на гнездо не делает. Но как только опасность для гнезда исчезает, птица возвращается на него почти сразу же. Такая тактика поведения является довольно эффективной и в сочетании с небольшими размерами гнезда и его довольно хорошей маскировкой приводит к тому, что гнездо не бросается в глаза и обнаружить его в лабиринте однотипных холмов без специальных навыков бывает довольно сложно.
Постэмбриональное развитие. Линька
Вылупившийся птенец покрыт густым коротким белым пухом (кроме голого участка брюха вокруг пуповины диаметром примерно 2,5 х 5 см и голого кольца вокруг глаз), на голове пух удлинен; клюв – желтоватый в основании и сильно темнеющий к окончанию, восковица голубоватая; лапы белые или розовые, цевка спереди опушена примерно только на 1/3, сзади – целиком голая. В выкармливании птенца активно участвуют оба родителя.
По наблюдениям за одним птенцом в гнезде установлено, что не раньше, чем на 10-й день неоптиль у него стал сменяться во второй пуховой наряд (который также чисто белого цвета) и одновременно с этим под пухом начали проклевываться колодки перьев; на 17-й день появились “кисточки” перьев (в том числе маховых и рулевых, скрытые еще под пухом; на 23-й день растущие опахала перьев уже начали покрывать пух, опахала маховых развернулись на 5-15 мм, надклювье у птенца начало темнеть, в промежутке между 17-м и 23-м днем отпал птенцовый зуб; в 50-дневном возрасте птенец уже был практически полностью оперен (до окончательной длины немного не доросли только маховые и рулевые перья) и достигал размера взрослой птицы, но еще не летал. Эти и другие наблюдения указывают, что в норме молодые становятся на крыло примерно в 2-месячном возрасте или чуть постарше, хотя в литературе для других районов указываются иногда и более поздние сроки – до 2,5-3 месяцев (Аверин, 1981; Митропольский и др., 1987; Рустамов, 1985; Makatsch, 1974).
Первые уже достаточно уверенно летающие сеголетки на Юго-Западном Копетдаге начинают встречаться в последней декаде июля (самая ранняя регистрация – 21.VII 1987, слеток с родителями парил над своим гнездом) – в первых числах августа. Семья и после вылета птенца может еще долго держаться в районе гнезда – иногда до конца сентября, т.е. до самого отлета.
Сроки линьки у змееяда практически не изучены, поэтому представляют интерес любые наблюдения на этот счет. 22.V 1992 мы наблюдали гнездящуюся пару, в которой у одной из птиц симметрично выпало несколько второстепенных маховых перьев, а у другой птицы – несколько второстепенных и первостепенных. 27.V 1995 под гнездом змееяда найдено три свежевыпавших первостепенных маховых пера, принадлежащих скорее всего самке.
Успешность размножения, случаи гибели
Гибель взрослой птицы отмечена нами только один раз: самка была убита каким-то хищником (скорее всего, полосатой гиеной Hyaena hyaena), видимо, прямо на гнезде, так как ее остатки валялись на этом же склоне всего в 20 м от жилого гнезда со свежими зелеными веточками. Гибель яиц установлена в трех случаях (18,8% из 16 гнезд с известной судьбой кладок), птенцов в гнездах – только в одном (10% из 10 гнезд с известной судьбой птенцов). Одна из кладок была разорена людьми, вторая – вероятнее всего гнездящимися поблизости воронами Corvus corax в момент отсутствия побеспокоенных людьми змееядов на своем гнезде, причины гибели двух остальных гнезд остались неизвестными. Расчетная успешность размножения, таким образом, составляет 0,73 вылетевших птенца на одну размножавшуюся пару, или 37-44 слетка в год на всю юго-западнокопетдагскую популяцию. Иногда птенцов из гнезд (так как последние очень доступны) забирают для забавы подростки – нам известен, по крайней мере, один такой факт; впоследствии этот птенец был выкормлен в заповеднике и в 1,5-годовалом возрасте успешно выпущен на свободу.
Трофические связи
По питанию змееяд – преимущественный герпетофаг. Специально его диету мы не изучали, но среди поедей в гнездах мы находили остатки кобры Naja oxiana, гюрзы Vipera lebetina, краснополосого Coluber rhodorhachis и оливкового С. najadum полозов, стрелы-змеи Psammophis lineolatum, желтопузика Ophisaurus apodus, шкурки ежей Hemiechinus sp. Змей птицы умерщвляют, перекусывая позвоночник в 2-3 см от головы. Слеток, содержавшийся в неволе, кроме рептилий и мышевидных грызунов, охотно поедал также озерных лягушек Rana ridibunda. Принесенных ему змей он сначала хватал лапой возле переднего конца (иногда двумя лапами сразу за оба конца), а затем расклевывал голову.
Нуждаются ли змееяды в постоянных источниках воды, осталось неясным, но на водопое мы их не наблюдали ни разу. Птенцы, не говоря уже о взрослых птицах, хорошо переносят жару. Маленьких птенцов родители закрывают на гнезде от солнца своей тенью. Подросшие же нередко основную часть времени, пока родители охотятся, проводят на солнцепеке. Во время сильной жары у птенцов вокруг ноздрей и клюва выступают кристаллики соли.
Охотятся на открытых участках, высматривая добычу сверху. При этом используются следующие основные способы: парение широкими кругами на большой высоте; поисковый облет на небольшой высоте над вершинами холмов с чередованием активного машущего полета и планирования; высматривание добычи с дающих хороший обзор присад (вершины холмов, столбы ЛЭП и т.п.). Могут подолгу зависать против ветра в воздухе на одном месте. Добытую жертву предпочитают съедать в определенном облюбованном месте на вершине какого-нибудь холма или скалы. Точки таких “столовых” (используемые часто и для ночевок) довольно постоянны и используются в разные годы. Добычу хватают на земле лапами, но переносят по воздуху в клюве (змеи при этом очень часто наполовину заглатываются вперед головой и из клюва свисает только хвост) и лишь как исключение – в лапах. Взрослые птицы и подросшие птенцы змей заглатывают целиком, маленьким птенцам скармливаются кусочки разорванной добычи.
Принятые и предлагаемые меры охраны
На территории Сюнт-Хасардагского заповедника (площадь около 300 км2) найдены гнезда трех и предполагаются гнездовые участки еще четырех пар, в том числе на Центральном участке заповедника гнездится три, на Чандырском – три пары и на Айдеринском – одна пара. Таким образом, под охраной находится примерно 12-14% местной гнездящейся популяции. В целях увеличения этого показателя очень перспективным является заповедание опустыненных низкогорий в районе слияния р. Чандыр с долиной Варказ (урочище Карахан), где представлены оптимальные местообитания змееяда и высока его гнездовая плотность.
Список литературы
Абдусалямов И.А., 1971. Фауна Таджикской ССР. Т. 19. Ч. 1. Птицы. Душанбе: Дониш. С. 1-403.
Аверин Ю.В., 1981. Отряд хищные птицы // Птицы (серия “Животный мир Молдавии”). Кишинев: Штиинца. С. 86-110.
Атаев К., 1974. Некоторые данные по экологии туркменского змееяда в восточных Каракумах // Материалы VI Всесоюз. орнитол. конф. Т. 2. М.: Изд-во МГУ. С. 19-20.
Бельская Г.С., Сухинин А.Н., Ташлиев А.О., 1966. Характеристика населения птиц некоторых районов Западного Копетдага. Сообщ. 3 // Изв. АН ТуркмССР. Сер. биол. наук. № 6. С. 54-59.
Воiнственський М.А., 1984. Птахи. Київ: Рад. школа. С. 1-304.
Гавриленко Н.И., 1929. Птицы Полтавщины. Полтава: Полтав. союз охотников. С. 1-133.
Дементьев Г.П., 1951. Отряд хищные птицы // Птицы Советского Союза. Т. 1. М.: Сов. наука. С. 70-341. – 1952. Птицы Туркменистана. Т. 1. Ашхабад: Изд-во АН ТуркмССР. С. 1-547.
Ефименко Н.Н., 1989. Редкие малоизученные виды птиц Копетдагского заповедника // Экологические аспекты изучения, практического использования и охраны птиц в горных экосистемах (тезисы докл. Всесоюз. симпозиума). Фрунзе: Илим. С. 34-36. – 1990. Современное распространение и динамика численности редких соколообразных птиц Центрального Копетдага // Редкие и малоизученные птицы Средней Азии (материалы III Республ. орнитол. конф.). Ташкент: Фан. С. 65-67. – 1990а. Многолетняя динамика численности соколообразных птиц на территории Центрального Копетдага // Заповедники СССР – их настоящее и будущее (тезисы докл. Всесоюз. конф.). Ч. 3 (зоологические исследования). Новгород. С. 230-233.
Зарудный НА., 1896. Орнитологическая фауна Закаспийского края (Северной Персии, Закаспийской области, Хивинского оазиса и равнинной Бухары) // Материалы к познанию фауны и флоры Российской империи. Отд. зоол. Т. 2. С. 1-555.
Ивановский В.В. 1992. Экология гнездования змееяда в Белорусском Поозерье // Современная орнитология. М : Наука. С. 69-77.
Корелов М.Н . 1962. Отряд Хищные птицы // Птицы Казахстана. Т. 2. Алма-Ата: Изд-во АН КазССР. С. 488-707.
Митропольский О.В., Фоттелер Э.Р., Третьяков Г.П.. 1987. Отряд Соколообразные // Птицы Узбекистана. Т. 1. Ташкент: Фан. С. 123-246.
Мищенко Ю.В., 1984. Орнитофауна Копетдага и ее зоогеографический анализ // Дисс. … канд. биол. наук. Киев. С. 1-505. – 1984а. Орнитофауна Копетдага и ее зоогеографический анализ // Автореф. дисс. … канд. биол. наук. Киев. С. 1-23. – 1986. Видовой состав и вертикально-биотопическое распределение птиц Центрального Копетдага // Природа Центрального Копетдага. Ашхабад: Ылым. С. 120-162.
Никифоров М.Е., Яминский Б.В., Щкляров Л.П., 1989. Птицы Белоруссии. Справочник-определитель гнезд и яиц. Минск: Выш. школа. С. 1-479.
Полозов С.А., 1982. Птицы Юго-Западного Копетдага (сезонная динамика состава авифауны) // Природа Западного Копетдага. Ашхабад: Ылым. С. 158-202. – 1983. Соколообразные Западного Копетдага // Экология хищных птиц (материалы I совещ. по экологии и охране хищных птиц). М.: Наука. С. 134-136.
Рустамов А.К., 1985. Птицы // Красная Книга Туркменской ССР. Т. 1. Позвоночные животные и высшие растения. Ашхабад: Туркменистан. С. 105-208.
Симакин Л.В.. 1989. Сведения по биологии туркестанского змееяда в Бадхызе // Изв. АН ТуркмССР. Сер. биол. наук. № 4. С. 69-70.
Скалон Н.В., 1982. Земноводные и пресмыкающиеся Юго-Западного Копетдага // Природа Западного Копетдага. Ашхабад: Ылым. С. 146-157.
Степанян Л.С. Степанян Е.Н., 1972. Материалы к орнитологической фауне Западного Копетдага // Фауна и экология животных. М.: Изд-во МГПИ. С. 193-213.
Сухинин А.Н., 1957. Материалы по экологии туркестанского змееяда в Бадхызе // Изв. АН ТуркмССР. № 5. С. 133-135.
Ташлиев А.О., Сухинин А.Н., Бельская Г.С., 1965. Характеристика населения птиц некоторых районов Западного Копетдага // Изв. АН ТуркмССР. Сер. биол. наук. № 4. С. 45-50.
Филатов А.К.. 1985. Материалы по экологии хищных птиц Западного Копетдага // Растительность и животный мир Западного Копетдага. Ашхабад: Ылым. С. 154-161.
Шерешевский Э.И., 1931. К биологии орла-змееяда в Крымском заповеднике // Сб. работ по изуч. фауны Крымск. заповеди. М.-Л.: Медгиз. С. 88.
Штегман Б.К.. 1937. Дневные хищники. Фауна СССР: Птицы. Т. 1. Вып. 5. Л.: Изд-во АН СССР. С. 1-294.
Щербак Н.Н., Хомустенко Ю.Д., Голубев М.Л., 1986. Земноводные и пресмыкающиеся Копетдагского госзаповедника и прилежащих к нему территорий // Природа Центрального Копетдага. Ашхабад: Ылым. С. 76-110.
Makatsch W., 1974. Die Eier der Vogel Europas. Bd. 1. Leipzig. S. 1-468.
Сюнт-Хасардагский государственный заповедник,
пос. Кара-Кала, Туркменистан
пос. Кара-Кала, Туркменистан
Поступила в редакцию 8 сентября 1994 г.
S.A. BUKREEV
On the biology of Circaetus gallicus heptnery in the south-western Kopetdag
Sjunt-Hasardag State Nature Reserve, Kara-Kala, Turkmenistan
The distribution and population density of Circaetus gallicus heptnery in the south-western Kopetdag are characterized basing on the literary data and the study carried out by author. The data on phynology, nesting and breeding success are given along with the descriptions of nests, eggs and postembryonal development of chicks. The measures taken to protect the species in the region are analyzed. Additional protective measures are proposed.